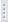Netýkavka žláznatá je statná jednoletá bylina, někdy až 3 m vysoká. Má přímou, světle zelenou větvenou lodyhu, která je zvenku tupě hranatá a uvnitř dutá. Dolní lodyžní listy jsou vstřícné nebo střídavé, horní vstřícné nebo v přeslenech, řapíkaté, vejčité až kopinaté a ostře pilovité. Květy jsou ve 2 až 20květých hroznech v úžlabí listenů, jsou červenofialové, růžové nebo až téměř bílé, uvnitř tmavě tečkované, s ostruhou a voní po ovoci. Kvete v srpnu až říjnu. Plodem je pukající tobolka vystřelující semena až do vzdálenosti 4 m. Jedna rostlina dokáže vyprodukovat až 5000 semen. Toto je obecně důvodem jejího rychlého šíření (Slavík, 1997).
Netýkavka žláznatá je považována v celé Evropě za invazní druh, i když ne tak agresivní jako např. křídlatky nebo bolševník. Nebezpečí netýkavky spočívá v tom, že mění složení rostlinných druhů v oblastech svého výskytu a díky své značné konkurenční zdatnosti vytlačuje původní druhy. Postupně tak dochází k přeměně původních rostlinných společenstev v druhově chudá společenstva s převahou netýkavky. Její vyplenění na některých plochách by bylo možné, i když ne snadné, ale existují obavy, aby to neotevřelo prostor pro ještě invazivnější rostliny (Hejda et al., 2009). Dalším invazním druhem netýkavky je v Evropě původem rovněž asijská netýkavka malokvětá (Impatiens parviflora) (Slavík, 1997).
Netýkavka žláznatá obsahuje ve svých nadzemních částech vedle dvou glukosylovaných steroidů, glanduliferinů A a B (Cimmino et al., 2016), zejména značné množství naftochinonů: 2-hydroxy-1,4-naftochinon a jeho 2-methoxy-derivát, jejichž obsah v rostlině se pohybuje kolem 0,8 až 1,1 %. To je mnohonásobně víc, než u ostatních netýkavek (Lobstein et al., 2001). Důvod, proč jsou invazní rostliny tak úspěšné v boji s původní vegetací, může mít několik příčin. Např. vyčerpání živin nebo stopových prvků, které pak ostatním méně agresivním rostlinám chybí (Dassonville et al., 2008). Důvodem ale může být také produkce specifických látek, syntetizovaných invazní rostlinou, které na ostatní působí jako herbicidy. Tyto alelochemikálie jsou fytotoxické a jsou odpovědné za to, že v nejbližším okolí takových rostlin žádné jiné rostliny nerostou (Rimando et al., 2003). Jako příklad je možno uvést česnáček lékařský (Alliaria petiolata). Před 150 lety se dostal z Evropy do Severní Ameriky a dnes na mnoha místech vytváří souvislé porosty, v kterých se už prakticky nevykytují žádné druhy původní americké flóry. V pozadí invazního úspěchu česnáčku je alelochemikálie sinigrin, který rostlina vylučuje svým kořenovým systémem do půdy a který ničí mykorhizní houby, žijící v symbióze s kořeny domácích rostlinných druhů (Lewis et al., 2006; Lankau, 2012). Naopak v Evropě je invazním druhem např. trnovník akát (Robinia pseudoacacia) původní v Severní Americe, který obsahuje látku robinetin (Nasir et al., 2005).
Fytotoxické alelochemikálie jsou po chemické stránce často látky s chinonovou strukturou. Takovým příkladem může být sorgoleon, lipofilní derivát p-benzochinonu, který je v podobě mikroskopických olejovitých kapiček vylučován kořeny čiroků (Sorghum spp.) do půdy (Patočka, 2013). Na každý plevelný druh, který by mohl ohrozit zdárný vývoj rostliny čiroku, působí sorgoleon jako velmi účinný herbicid (Weidenhamer, 2005; Dayan, 2006; Dayan et al, 2009). Skutečnost, že netýkavka žláznatá je producentem naftochinonů s možným bioherbicidním účinkem, je také možnou příčinou invazních vlastností této byliny.
Poslední výzkumy ukazují, že 2-hydroxy-1,4-naftochinon není nativní látkou. Tou je zřejmě 1,2,4-trihydroxynaftalen-1-O-glukosid a při analýze drogy nalezený 2-hydroxy-1,4-naftochinon může být produktem jeho odbourávání (Tříska et al., 2013).
Literatura
Cimmino A, Mathieu V, Evidente M, Ferderin M, Banuls LMY, Masi M et al. Glanduliferins A and B, two new glucosylated steroids from Impatiens glandulifera, with in vitro growth inhibitory activity in human cancer cells. Fitoterapia, 2016; 109: 138-145.
Dassonville N, Vanderhoeven S, Vanparys V, Hayez M, Gruber W, Meerts P. Impacts of alien invasive plants on soil nutrients are correlated with initial site conditions in NW Europe. Oecologia. 2008; 157(1): 131-140.
Dayan FE. Factors modulating the levels of the allelochemical sorgoleone in Sorghum bicolor. Planta 2006; 224: 339–346.
Dayan FE, Weidenhamer JD, Howell J. Dynamic root exudation of sorgoleone and its in planta mechanism of action. J Exp Botany. 2009; 60(7): 2107-2117.
Hejda M, Pyšek P, Jarošík V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J Ecology 2009; 97: 393–403.
Lobstein A, Brenne X, Feist E, Metz N, Weniger B, Anton R. Quantitative determination of naphthoquinones of Impatiens species. Phytochem Anal. 2001; 12(3): 202-205.
Lankau RA. Coevolution between invasive and native plants driven by chemical competition and soil biota. Proc Natl Acad Sci USA. 2012; 109(28): 11240-11245.
Lewis KC, Bazzaz FA, Liao Q, Orians CM. Geographic patterns of herbivory and resource allocation to defense, growth, and reproduction in an invasive biennial, Alliaria petiolata. Oecologia. 2006; 148(3): 384-395.
Nasir H, Iqbal Z, Hiradate S, Fujii Y. Allelopathic potential of Robinia pseudo-acacia L. J Chem Ecol. 2005 Sep;31(9):2179-92.
Patočka J. Sorgoleon, bioherbicid čiroku. TOXICOLOGY, 2013. Dostupné na http://toxicology.cz/modules.php?name=News&file=article&sid=590
Rimando AM, Dayan FE, Streibig JC. PSII inhibitory activity of resorcinolic lipids from Sorghum bicolor. J Nat Prod. 2003; 66: 42–45.
Slavík B. (1997): Impatiens L. – netýkavka. – In: Slavík B., Chrtek J. jun. & Tomšovic P. (eds), Květena České republiky 5, p. 230–240, Academia, Praha.
Tříska J, Vrchotová N, Sýkora J, Moos M. Separation and Identification of 1,2,4-Trihydroxynaphthalene-1-O-glucoside in Impatiens glandulifera Royle. Molecules. 2013; 18(7): 8429-8439.
Weidenhamer J. Biomimetic measurement of allelochemical dynamics in the rhizosphere. J Chem Enol. 2005; 31: 221–236.